NOTE: The logic of this draft remains unchanged,
but it contains two (approximately canceling) errors that will be corrected
in the next draft. 1) The calculations are in cgs-georgi units, rather
than mks-georgi units. The cgs-georgi unit system is
not CONSISTENT at the level of operational procedures, as that mks-georgi
system is. On some crossterms, this makes a difference.
2) The capacitance effect of the fluid clefts surrounding the neural lines
is not accounted for. The effects of 1) and 2) about
cancel. All the logic below stands unchanged.
******************************************************************************************************
HYPOTHESIS: DENDRITES,
DENDRITIC SPINES, AND
STEREOCILIA HAVE RESONANT MODES UNDER S-K THEORY
M. Robert Showalter(1)
The Showalter-Kline (S-K) equations for passive neural conduction predict
that in low g mode:
The stereocilia in the auditory inner hair cells are
sharply tuned resonant columns.
The dendritic spines in a channel-closed condition are sharply tuned passive
resonators that may have both a LRC and a column resonant mode. Opening
of one or a few channels shuts off spine resonant receptivity.
The dendritic branches, subbranches, or sections can, with spines, form
resonant geometries, and these geometries may be shaped and switched by
open synapses acting as impedance mismatch reflection sites.
In high g mode, spines and dendrites have very high dispersion similar
to that familiar under the Kelvin-Rall (K-R) equation, and have practically
no resonant properties.
Resonance
is logically interesting. Resonant systems can effectively
amplify signals of a tightly defined period and phase. Radio
and television offer familiar examples of resonant selectivity. Radio
and television receivers exist in an electromagnetic field consisting of
a bewildering and undescribable variety of electromagnetic fluctuations.
Reception occurs because the resonant receiver is selective
for a specific frequency at a high degree of phase coherence.
Signals off frequency are not significantly detected, and "signals"
of random phase that are on frequency cancel rather than magnify in resonance.
Radar receivers also operate on the principle of resonance.
Other examples are our telephone system and cable television system, each
organized so that a multiplicity of different signals can be carried in
physically mixed form over the same conduits. These "mixed"
signals can be separated and detected with negligible crosstalk by resonant
means. Under the Showalter-Kline (S-K) passive
neural conduction theory, dendritic spines, dendrites, and stereocilia
appear to be passive resonant elements adapted to tuning and sharp on-off
switching. These are interesting characteristics for these
components to have, for brains are organs of logical manipulation, and
oscillations in brain are common, complicated, and known to carry information(2)
(3) (4) (5).
Under the S-K theory, brain
communication occurs in TWO modes, by conduction and inductive resonant
coupling, not only by conduction. When dendritic sections
or spines are in the high transmission mode of S-K, neurons not
directly connected can "communicate" by inductive coupling over
the extracellular medium. Interesting possibilities follow
from this. Consider the set of logical interactions that can occur among
a population of people, all able to both broadcast and receive radio signals,
with that population operating over many wavelengths.
The number of interactive combinations in such an interacting set is prodigious,
and the coordinations possible very detailed. Under S-K,
much of the brain looks analogously adapted for sending and receiving signals
in the frequency domain. Frequency selectivity of dendritic
spines and spine assemblies is high (with predicted bandwidths often less
than .001 Hz and predicted Q's above 1000). Under
S-K, information flow in brain by inductive means may occur with
little or no crosstalk between many frequency-defined "bands"
(or multifrequency-time signatures, analogous to auditory words).
Under these conditions, the combinatorial limitation of "connectionist"
neural nets (due to the explosion of N! as N increases) goes away.
Because resonant communication and processing is inherently a massively
parallel process, the processing limitations of Feldman's "100 step
rule(6)" are also much softened.
For example, resonant processing appears to be capable of handling frequency
coded "list reading" functions, such as the lexical access problem,
in massive parallel, without need for the very time consuming sequential
search process. (For instance, searching one word out
of 50,000 might be one "resonant signal interrogation step,"
comprising the sending out a frequency-time signature of the word and its
resonant recognition by only one tuned recieving neuron among many.)
A major reason for rejecting "symbol processing AI," in spite
of its achievements and the inherent plausability of its symbol-processing
approach, has been the inherent slowness and "anatomical implausibility"
of the list-reading steps that AI algorithms often contain.
If neurons are capable of BOTH communication via conduction AND communication
by inductive resonant coupling, as S-K and anatomy together suggest,
some "theoretically impossible" animal accomplishments become
less mysterious.
Resonance-like behavior in brain has long
been observed. Evoked electroencephalographic data from
human and animal subjects analyzed by David Regan's zoom fast fourier transform
analysis(7) shows stunningly sharp resonance-like
behavior. Most of the EEG or MEG signal power is concentrated
at peaks that are exact integer multiples (or sums and differences) of
evoked stimulus frequency(s). The bandwidths of these peaks
are too tight to measure, perhaps less than .002 Hz. This tightness
is astonishing when one considers that EEG must measure integrated effects
over substantial volumes of brain. If one looked at these curves
from an engineering perspective, in ignorance of where they came from,
one would be strongly tempted to infer an ensemble of sharply coupled very
high Q passive resonators. When the S-K equations
of neural conduction are applied to anatomy, one gets an ensemble of high
enough Q resonators (dendritic spines) interacting via the extracellular
media, that should produce the results Regan measured.
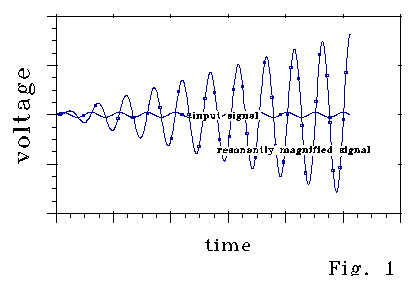
Fig 1 shows how voltage in a passive resonator
varies with input "signal" voltage. The perturbation
(input) signal is phased so that resonance builds up. If, at
some time, input signal phase shifted 180 degrees, the resonant signal
would decrease at the same rate. Around null, a shift
of a few degrees makes the difference between rather rapid resonant signal
increase and rapid decrease. FM radio operates on this principle.
Phase sensitivities and frequency selectivities of such passive resonant
systems are well exploited throughout communication technology.
Passive resonance seems like it may be adapted to similar sensitivities
long observed in intact animals, and now seen in neurons(8).
Passive resonant systems perturbed at exactly
their resonant frequency can build up energy to a value many times the
amplitude of the exciting disturbance. The steady state
resonant amplification factor, Q, is an important figure of merit
for passive resonant arrangements:

Bandwidth (the frequency difference between the half power points on a
resonance curve) is inversely related to Q according to the formula:
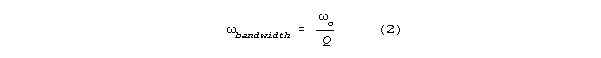
where o is resonant frequency. Resonant systems with low damping
have high Q's and tight bandwidths.
A transmission line terminated in pure capacitance
or an open or short circuit condition can have impedance properties identical
to those of lumped element, and can also act as a column resonant element.
Properly terminated, a transmission line may carry waves that repeat exactly
in a specific period, regardless of wave shape complexity, in the
same fashion that the air column of a clarinet will carry and summate extremely
complicated (but exactly periodic) waveforms(9).
Such lines or columns "store" repeating wave patterns that fit
the geometry of the line between reflecting ends. They
can have very high Q's. Transmission lines of 1/4 and
1/2 wavelength have been used as resonators in the communication fields
for nearly a century, and acoustically analogous columns have been used
in musical instruments for millennia. A good column resonantor
would store and summate exactly periodic patterns such as those of action
potential spikes, and operating in that mode on such signals could act
as a very precise timing or timing comparator component.
In engineering practice, the resonant
effects produced by electrical transmission line elements tend to happen
at very high frequencies ( >107 Hertz).
Under neural conditions as predicted by S-K, functionally identical
and strong resonant effects occur at much lower frequencies - frequencies
ranging from about 1 Hz to the frequencies of bat audition, above 100,000
hz.
A passive resonant system excited for enough
cycles at its resonant frequency by a signal of RMS voltage v will build
up oscillations to an RMS voltage of Qov.
Voltage buildup is rapid: after only five cycles of excitation, resonant
voltage amplification of fifteen or more times peak voltage occurs in a
high Q passive resonant circuit.
HAIR CELL STEREOCILIA AS 1/4 WAVE AND 1/2 WAVE RESONANT LINES:
Although an enormous amount of experimental
work has been done on the hair cells, the primary auditory sensors in animals,
our knowledge of how hair cells work remains incomplete and tentative.
Current explanations of hair cell frequency response may be
considered strained around about 400 Hz. Bat hair cells, which,
despite specializations, look much like other hair cells, function as precision
navigational input instruments at frequencies more than 2000 times higher.
When the S-K equation is used, the freestanding stereocilia of the
inner hair cells appear to be resonant transmission lines with the necessary
resonant frequencies. The idea that stereocilia are resonant columns may
be part of the explanation needed to explain the performance of real hair
cells that operate at usual and high mammalian and avian frequencies. The
Regans have measured signals that appear to occur at the hair cell level,
that show resonantly organized peaks consistent with column resonance in
the hair cells(10).
Well terminated lengths of neural passage
that are sharply open (short circuited) at 1/4 of wavelength L will
be resonant at the frequency omega corresponding to L and at integer
multiples of omega. An open stereocilium, or a stereocilium
with an open end channel, would be an example of such a resonant structure.
A well terminated length of neural passage that is one half wavelength
L, long and sharply closed on one end will also be resonant at frequency
and integer multiples of omega. A stereocilium terminated by
a closed channel would be an example of such a resonant structure.
It is worth noting that the primary and overtone resonant frequencies of
the open channel line are also resonant frequencies for the same stereocilium
in the closed end channel configuration.
Consider a stereocilium to be cylindrical,
with diameter d cm, membrane thickness cm, membrane dielectric factor
epsilon, central fluid conductance sigma ohm-cm, with a resonant frequency
omegar radians/sec and a corresponding resonant wavelength
L cm.
According to S-K theory(11), the
transmission velocity in the low g regime is
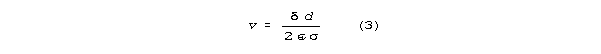
and the wavelength corresponding to frequency is
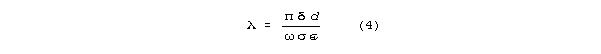
In this regime, Q>>1 and Q is inverse with attenuation
per wavelength .
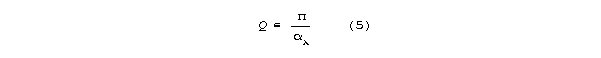
Substituting into (5) for a passage having 100% positive
or negative end reflection, Q is:
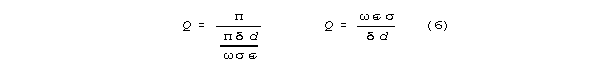
Bandwidth is
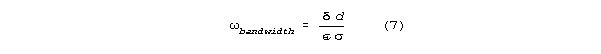
The column-resonant bandwidth for set diameter, conductance,
and capacitance is independent of resonant frequency (length).
Consider model stereocilia for set sigma = 110 ohm/cm
and
capacitance = 10-6 farads/cm2 for different
diameters.
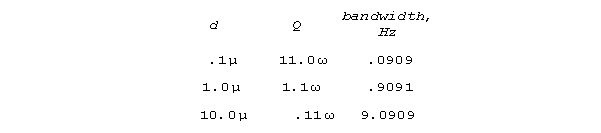
For these conditions and a resonant frequency of 1000 rads/sec (159.15
Hz) a free standing and well terminated .1 micron stereocilium will have
a Q of 11,000 and a bandwidth of .0909 Hz. (A stereocilium in a
bundle of other stereocilia would be more complicated.) As
a 1/4 wave line, length for this freestanding stereocilium would be 1.571
microns; as a 1/2 wave line, length would be 3.142 microns. Lower
frequencies would correspond to some combination of longer stereocilia,
larger , or smaller diameter. Higher frequencies would
require increased diameter; reduced capacitance (for instance, by increasing
stereocilium wall thickness, as is often observed), packing stereocilia
together; smaller resistivity , or some combination. Resonant performance
degrades as passages become larger. Q is proportional to resonant
frequency (inverse with length). At bat frequencies, extremely high Q's,
very well adapted for information processing, are predicted by this simple
S-R theory.
Measurements at these scales are not yet
possible, and estimates of values are therefore speculative. Even
so, on the basis of microanatomy and chemistry it seems reasonable to expect
properties adequate for strong 1/4 and 1/2 wave resonance in stereocilia
at frequencies ranging from 106 Hz (in bats) down to a few Hz.
Q's of 103-106 would be expected with essentially
perfect end reflection. The stereocilia in inner hair
cells are geometrically arranged in a fashion to maximize impedance mismatch
end reflection.
Under S-K theory, stereocilium bundling
in inner hair cells would seem to be an adaptation to prevent stereocilium
destruction. Under S-K theory, a single high Q
stereocilium would be a fragile and short-lived structure in an ordinary
cochlea. An input signal might easily build up to a voltage
greater than the dielectric breakdown strength of the stereocilium wall,
destroying the stereocilium. An adaptively changeable stereocilium
length would reduce this kind of fragility, but rather large length changes
would be needed to protect single stereocilia against strong, long duration
signals. High sensitivity and stereocilium survival would therefore
tend to be incompatible for single stereocilia. (Except for
stereocilia in inner hair cells adapted for a short, stereotyped duty cycle.
Bat inner hair cells, that have such duty cycles, have stereocilia "bundles"
with single or a few stereocilia(12) .)
Bundling of stereocilia
capable of some length change would offer a defense against such resonant
destruction. If there were two or more stereocilia connected
to a single cavity, and if the stereocilia had even a small ability to
change length, high sensitivity would be compatible with structural survival.
If input sound stimulus was too high, a small shift in length of one or
more of the stereocilia would reduce the Q of the stereocilia assembly
by destructive interference. Length changes of 10-3
or less would produce strong attenuation in a previously high Q
system. Assembly Q might be rapidly changed from
104 to 10 (or vice versa) with such an arrangement. Stereocilia
contain contractile proteins that may be adapted and somehow controlled
for this purpose.
The inner hair cell is a complex structure
in a complicated environment. It includes ornate mechanical
linkage, stereocilia end channels that open and close on the basis of mechanical
stimuli and would appear to shift stereocilia between 1/4 wave and 1/2
wave mode, structurally ornate arrangements of the stereocilia, and sensitive
and almost certainly temporally tuned channel and messenger kinetics in
the hair cell body. Detailed discussion of these important
matters is beyond the scope of this paper. However, the high
Q characteristics of the stereocilia interpreted according to the S-K
equation fit well with what we know that hair cells do.
Resonant effects
also appear likely in the stereocilia of the outer hair cells, and may
be involved in their membrane tension controlling functions.
DENDRITIC SPINE RESONANCE:
Under S-K theory,
combined with the assumption that the spines can exist in an all-channel-off
condition, the spines are predicted to have a very high Q passive
resonant mode. From an information processing perspective,
this should be an infrequent mode, with the more common mode very much
like the high dissipation mode(13) widely
assumed. The high resonance mode requires that all spine channels
be off, and may perhaps be easily turned off in experimental manipulation.
The resonant mode description here is no complete picture of spine function,
but it is believed to be consistent with what is now known about spines.
Fig 2(14)
shows camera lucida drawings of a neuron body and proximal dendrites of
a neuron showing common spine types: thin, mushroom shaped, and stubby.
Aspect ratios and sizes of all these types vary.
A stubby type with a thinner aspect would be subject to the same logic
described for a closed end stereocilia. The "thin"
type and "mushroom" type can be considered as both an LRC element
and a column resonant elements. In LRC (inductance-resistance-capacitance)
mode, the bag and shaft have a lumped capacitance, and shaft inductance
and resistance are considered in lumped form. In column mode,
the shaft is a column open at both ends, with a capacitive termination
correction at each end.
Fig. 2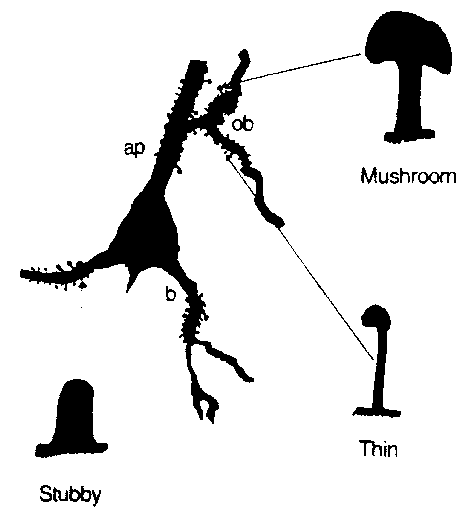
Fig. 3 shows an electrical model of a thin spine. In the
scaled figure, 4/5 of spine capacitance is in the bag section.
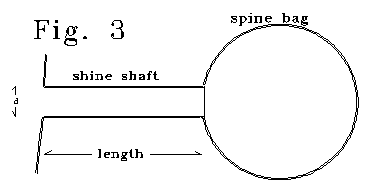
The "spine" of Fig. 3 can be modelled as an LRC resonant system.
Capacitance is the capacitance of the "bag" section, plus half
the capacitance of the shaft section. The shaft has resistance of R, and
an effective inductance of Lex. The Le/R ratio is
inverse with diameter. Different bag sizes for the same shaft size yield
different LC products, and different resonant frequencies. Q, radian
frequency, and bandwidth, for the LRC case are:

In the model, bandwidth is proportional to diameter, as in
the stereocilia case. Let's arbitrarily chose a shaft diameter
of .1 µ, shaft length of .5 µ, interspine medium
conductance of 110 ohm-cm, membrane capacitance of 1 microfarad/cm2,
and zero membrane leakage, g. Holding these values,
and varying bag size, yields the following relation between frequency and
Q.
Q = o(910) (9)
For Regan's measured frequency range of 7-45 hz (44-283 radians/sec,)
Q's between 40,000 and 257,000 are estimated. Regan's
data correspond to Q's about a decade smaller, between 3,680 and
24,000 over that same 7-45 Hz frequency range. This is an acceptable fit
because:
Regan must have measured ensemble properties, not properties of single
neural elements.
Regan's setup could have detected no tighter bandwidths than he did detect.
and
Within the constraints of biological knowledge, we could have guessed other
values of the parameters to come closer to Regan's values (or even to match
them.)
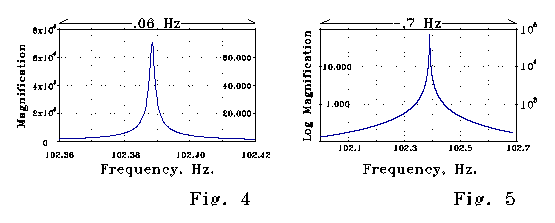
Figs 4 and 5 below show steady-state
magnification of a signal as a function of frequency calculated for the
LRC spine model of Fig 3. The peak magnification factor is
about 70,000. Note the sharpness of the magnification as a function of
frequency.
The model spine of Fig 3 would also have a column resonance
mode.
Spine column resonant frequency will be approximately

column resonant Q from equation (6) ***** check deriv** is
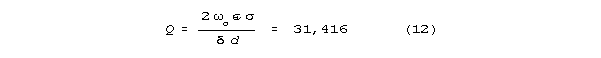
Electrical compliance of the bag would shift these resonant frequencies
and Q's somewhat from the simple 1/4 wave column calculation set
out above, but the correction would involve details that can be considered
elsewhere.
Referring again to Regan's
data, the brain has more than 1013 spines. If spine
resonant frequencies are widely distributed in brain, and some fraction
(say .001) of the dendritic spines are in the high Q state, one
would expect long duration fixed frequency stimuli, such as Regan supplied,
to yield the sort of excitation curves that Regan observed.
Coupling of the spines would be via the very rapid conduction of
the extracellular medium, not via conduction along dendrites or axons.
SPINE SWITCHING:
According to S-K,
opening of one channel will change a spine from a sharply tuned state,
with a Q in the thousands, to an untuned Q of 10 or less.
A single channel therefore acts to switch spine receptivity on or off.
This may be useful for neural logic, and also to eliminate
resonant overload spine destruction analogous to the stereocilia destruction
discussed before. (I believe that the extensive destruction of spines and
dendrites that occurs in severe epilepsy(15)
may happen in this way.) Suppose there is one membrane
channel open in the bag portion of the spine. That channel
will act as a resistive, energy dissipating shunt. The spine
will have a shunt limited Q, Qdamped, that is
much less than the undamped Q. Let Rc
be shunt channel resistance. If Qdamped<<Q
(which makes sense for reasonable channel conductances (between 4 and 400
picosiemens) then
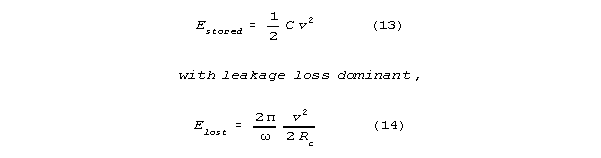
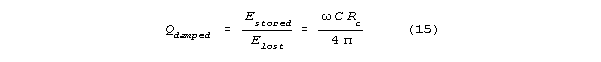
IMPEDANCE SWITCHING:
Under S-K, dendritic conduction in
the low g regime can be sharply damped, and reflected, by synapses
that change local values of g quickly. Impedance
jumps in neural passages due to changes in g can act analogously
to similar sharp impedance jumps that occur at pads or finger holes in
woodwind musical instruments. If the change is relatively
large, this reflection can be both sharp and strong.
Reflecting discontinuities can be set up by synaptic or dendritic spine
activity. Neurons appear to be abruptly switchable impedance
mismatch devices capable of switching whole dendritic trees or branches
on and off. (Such trees or branches may contain hundreds,
or thousands, or even hundreds of thousands of potentially resonant spines.)
Under S-K, one can therefore speak
of dendrites, with spines, between open synapses, as more-or-less complicated
"electroacoustical" resonant assemblies capable of resonating
to patterns considerably more complicated (and more information-bearing)
than sine waves. My initial calculations indicate that time-frequency
patterns as complicated as phonemes and simple words may be possible for
anatomically plausible resonant assemblies with simple switching under
S-K, and I have become interested in how such patterns might be
learned, sent, and recieved by inductively coupled neuron-like systems.
An examination of the anatomy
of pyramidal cells, basket cells, chandelier cells and other neural types
shows a multiplicity of axodendritic synapses at the interfaces between
axons and cell bodies, and at many branch points in the dendritic arborization.
These synapses are specific to cell type and geometrically ornate. According
to the S-K theory these synaptic structures would function to sharply
"switch" specific portions of dendritic anatomy (which may contain
thousands of spines) into or out of effective contact with the cell body
and axon hillock by production of sharp impedance mismatch reflections.
This theoretical prediction is totally unproved. However, if
the prediction is true, it greatly increases the amount of information
processing possible in a single neuron or small group of neurons, compared
to the maximum possible under K-R.
According to the hypothesis set
out here, neural function has much in common with the function of electronic
communication circuits. Even so, there are pronounced differences:
The lines themselves are bistable.
Conduction along neural lines occurs in two very different modes, depending
on the value of g determined by channel openings.
Neural line inductance is inverse with the cube of diameter, where electronic
line inductance is diameter-independent in the absence of magnetic inductance.
This means that a spiny dendrite is no simple analog of an antenna, even
without the complicated switching that exists according to S-K.
Neurons have huge numbers of many widely distributed and variously controlled
current sources (ion channels). and
Under S-K, neurons are capable
of prodigious numbers of rapidly variable switching states.
One theoretical implication of S-K theory is this:
resolution of much neural function will have to be determined by experiment.
There are too many combinations to conceive of or choose
from. However, theory may sometimes be useful to identify
patterns and choose among alternatives.
When the S-K equations for passive neural conduction
are substituted for the K-R equations, resonant and switching mechanisms
that appear adapted to audition and neural logic follow from simple calculations
combined with our knowledge of neural anatomy.
The S-K equations themselves
follow from a conclusion that can be expressed in two ways:
1) J.C. Maxwell's first method is the proper method for deriving complete
equations from physical models(16) or,
equivalently
2) In derivations from physical models, crosseffect terms
involving dimensional parameters must be algebraically simplified in intensive
form(17).
These alternative statements of
a conclusion in the theory of physical modelling are therefore of interest
in neurobiology.
NOTES
1. Department of Curriculum and Instruction, School
of Education, University of Wisconsin, Madison USA email: showalte@macc.wisc.edu
2. Llinas, R. Intrinsic electrical properties
of nerve cells and their role in network oscillation Cold Spring
Harbor Symposia on Quantitative Biology 55, 933-938 (1990).
3. Llinas, R., Ribary, U. PNAS 90 2078-81
(1993).
4. Wehr M. & Laurent, G. NATURE 384,
162-166 (1996).
5. Naatanen R. et al NATURE, 385 432-434
(1997).
6. Feldman, J.A. COGNITIVE SCIENCE 9,
1-2 (1985).
7. David Regan HUMAN BRAIN ELECTROPHYSIOLOGY:
Evoked Potentials and Evoked Magnetic Fields in Science and Medicine
Elsevior pp. 106-108 (1989).
8. Koch C. NATURE 385 207-210 (1997).
9. Newman, R.B SOUND, ENCYCLOPEDIA BRITANNICA
15 Figs 7, 8.
10. Regan, op. cit. pp 276-278.
11. Showalter, M.R. A New Passive Neural Equation
Part a: derivation. available FTP angus.macc.wisc.edu/pub2/showalt
12. Vater M, Lenoir M. J of COMP. NEUROLOGY
318(4) 367-79 (1992).
13. Rose, G.J., Call, S.J. PNAS 89
9662-5 (1992).
14. C.H. Horner Plasticity of the Dendritic Spine,
Progress in Neurobiology, v.41, 1993 pp. 281-231, Fig 3,
p 285.
15. Muller, M., Gahwiler, B.H., Rietschin, L. PNAS
90 257-61 (1993).
16. Showalter, M.R. and Kline, S.J. Modeling
of Physical Systems According to Maxwell's First Method. available
FTP
angus.macc.wisc.edu/pub2/showalt
17. Showalter, M.R. and Kline, S.J. Equations
from Coupled Finite Increment Models Must be Simplified in Intensive Form
available FTP. angus.macc.wisc.edu/pub2/showalt